

 August 25th, 2012
August 25th, 2012  riffin
riffin INTRODUCTION
Some people think dinosaurs were cold-blooded; other people think dinosaurs were warm-blooded. Lots of scientists have studied the problem. There is information about posture, bone structure, and brain/body size. This essay will consider the evidence and try to come to a reasonable conclusion.
THE BIG DEBATE
They were reptiles, so they must have been cold-blooded
In the old days, before about 1960, everybody thought dinosaurs were cold-blooded, that is, their temperature was determined by that of the external environment. Scientists thought this because they had decided that dinosaurs were reptiles, and all modern reptiles are cold-blooded. (It should be noted that the term “reptile” has fallen out of favor in the modern, cladistic view, denoting as it does a paraphyletic group; however, most people still understand what is meant by a reptile, and I will use the term throughout in its traditional sense, making no pretense that it designates a natural clade.) Alligators and crocodiles are the most closely related non-dinosaurian archosaurs, and they are cold-blooded.
Then people began to look further, and some of them decided that it was not so easy. Dinosaurs were unusual reptiles at best. They do not look like modern reptiles. They did not walk like modern reptiles. It had been suggested almost a century ago that birds were the direct descendants of dinosaurs, and they are indisputably warm-blooded; in fact, in the cladistic view, birds are dinosaurs. Maybe there was more to the story.
They held the heat because they were so big

stegosaurus
Several scientists proposed the “inertial homeotherm” idea. They said a big dinosaur could keep an even body temperature just because it was so big. According to Colbert and colleagues, a medium-sized dinosaur exposed to day and night temperatures similar to those found in Florida today would take 86 hours to change its temperature by 1°C. They based their estimates on experiments with alligators tied up out in the sun. Mesozoic climates were far more equable than they are now. Clearly, by moving from sun to shade, as modern ectotherms do, a fine degree of temperature control could be achieved without invoking a high metabolism.
They might have been warm-blooded
Postural evidence
Bakker was the first scientist to come out strongly in favor of warm-blooded dinosaurs. He had several arguments. He noticed that dinosaurs all had an upright posture, instead of a sprawling one like modern reptiles. Bakker divided today’s animals into three groups, based on their stance.
Group 1 is the sprawlers. They have their bellies on the ground most of the time. This group includes salamanders, frogs, snakes, lizards, turtles, the tuatara, and monotremes.
Group 2 is the semi-erect animals. They lie down on their bellies, except when they want to go somewhere. Then they walk pretty much upright. This group includes chameleons, alligators, and crocodiles.
Group 3 is the fully erect animals. They usually stand up unless they are resting. This group includes birds, marsupials, and placental mammals.
Bakker noticed that the fully erect animals were all warm-blooded. Monotremes are warm-blooded even though they sprawl. He couldn’t prove that only warm-blooded animals could be fully erect, but it was an interesting correspondence.
Bakker said that fully erect animals spend more time moving around than sprawlers do. Lizards can raise their metabolism by moving around . Bakker proposed that the evolution of erect posture was connected to activity level and warm-bloodedness.
Monotremes are thought to be the most primitive living mammals. Their resting metabolic rate is lower than other mammals. Their body temperature is also lower than that of other mammals. Obviously, an animal can be a sprawler and still be warm-blooded. But can a cold-blooded animal maintain a fully erect posture?
Bakker’s critics say yes. They point out that upright posture is more efficient, and big animals need to stand upright to support their large weight. (Then why could Permo-Triassic rhynchosaurs and parieasaurs grow to the size of rhinos and still retain a sprawling posture?). The critics say Bakker has not proven a causal relationship between posture and body temperature. But then, it’s pretty hard to prove a causal relationship in paleontology. You can’t do any experiments on the animals you are investigating.
Bone histology
A couple of years later, some new evidence came to light. De Ricqles studied the structure of fossil and recent bones under the microscope. He noticed that the bones of modern warm-blooded animals had a lot more Haversian canals than the bones of cold-blooded ones. The cold-blooded animals’ bones also had growth rings in them, forming what he called a “lamellar-zonal” pattern. He called the warm-blooded type “fibro-lamellar” structure.
De Ricqles said that the distinction between the two types was really the effect of growth rate. Fast-growing animals have to remodel their bones more extensively than slow-growing ones. Based on the bones of living animals, he concluded that only warm-blooded ones had a high enough metabolism to support the fast growth leading to fibro-lamellar bone.
The group of diapsid reptiles called archosaurs includes the ornithischian and saurischian dinosaurs, and also pterosaurs and crocodilians. De Ricqles found fibro-lamellar bone in the first two groups. The dinosaurs he examined included carnosaurs, prosauropods, sauropods, and ornithischians. He found lamellar-zonal bone only in crocodilians. He also found evidence of fibro-lamellar bone in the advanced thecodonts ancestral to the first three groups of archosaurs, and in the advanced therapsids ancestral to mammals. Bakker and Feduccia both suggested perhaps endothermy arose as a response to the rigorous climates of the Permian southern hemisphere glaciations.
De Ricqles notes that warm-bloodedness need not denote a body temperature of 37°C. Monotremes maintain about 28° to 30°C. Marsupials and primitive placentals like hedgehogs and tenrecs have metabolic rates about 3/4 those of other placental mammals and body temperatures 3° to 5°C lower. It is also possible that the degree of warm-bloodedness varied among dinosaur groups. It has recently been suggested that some dinosaurs may have had high metabolisms and growth rates while young, slowing to become inertial homeotherms as they reached adult size.
Bouvier claims that slow-growing mammals like monotremes and bats have poorly vascularized bone. However, this does not negate the value of Haversian bone in diagnosing warm-bloodedness.
Insulation
Everyone seems to agree there is no need to insulate a cold-blooded animal. It keeps the same temperature as its environment and doesn’t care. Fossils of pterosaurs have been found with hairlike structures on the skin. Besides, scientists believe an animal must be warm-blooded to support the activity of flight. So they do not argue too much about pterosaurs having been warm-blooded.
The earliest widely acknowledged bird is called Archaeopteryx. It is from the Jurassic Period, about 140 million years ago. It had feathers, but a skeleton indistinguishable from a small dinosaur called Compsognathus. So it is a matter of semantics whether it is called the first bird or a feathered dinosaur. Recently, another, slightly younger dinosaur fossil was unearthed in China that sported a row of proto-feathers down its spine, and dinosaurs with indisputable feathers attached to the forelimbs were described in 1998. These finds have convinced scientists that the first birds evolved from small, meat-eating dinosaurs. Warm-bloodedness must have arisen in this lineage sometime prior to the appearance of Archaeopteryx, since it was insulated with feathers. Who is to say exactly how early on? Were there feathered tyrannosaurs?
Other dinosaur fossils do not bear feathers or fur. In some cases, it may be because delicate structures do not preserve well. Most fossils of mammals don’t show fur either. In other cases, skin impressions are known, and they are of a scaly, reptilian type. This doesn’t mean there can’t be smooth-skinned endotherms, though. Bakker points to hippos and elephants. They live in warm climates maybe similar to those in the Mesozoic, and don’t need fur to keep themselves warm. Aquatic mammals like whales live in cold environments and use blubber for insulation, leaving the skin smooth. There has been some suggestion recently of fat storage structures in dinosaurs. Insulation is not as important in big, warm-blooded animals as in small ones because of the difference in surface area-to-volume ratios.
Arctic and Antarctic dinosaurs
In the past decade, evidence has come to light regarding the existence of dinosaurs inside the Antarctic and Arctic circles). Fossils have been found in parts of Australia, Antarctica, and Alaska known to have experienced at least three months of annual darkness during the Cretaceous Period. Temperatures are thought to have hovered only slightly above the freezing point during the winter. The Alaskan hadrosaur herds included juveniles without much thermal inertia and probably without the capability of undertaking long migrations, yet they made it through the winter all right, possibly by hibernation. This is far from proof of warm-bloodedness in those dinosaurs, but it is suggestive.
BRAIN/BODY RELATIONSHIPS
Although it is less direct, there is another piece of evidence pertinent to the ectothermic vs. endothermic debate. This is the relationship between brain size and body size.
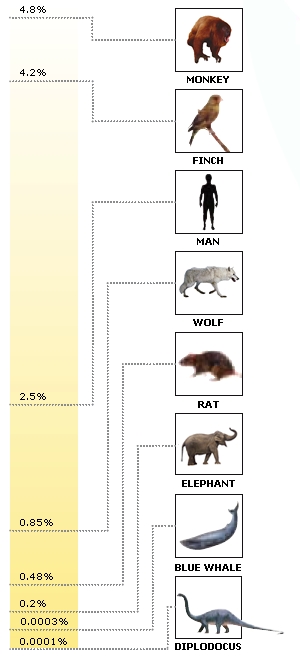
brain body relation of dinosaurs
It is often said that an animal’s brain size is a function of its body size according to the allometric equation
ya = b (x )
This means that when the logarithm of brain size is plotted on the y axis, and the log of body size is plotted on the x axis, you get a straight line. Linear regression is frequently used to fit the straight line to the data.
The surface-to-volume model
Jerison said the regression lines for lower and higher vertebrates have a slope of 2/3. Surface area is a function of linear dimension squared, and volume is a function of linear dimension cubed. Jerison speculated that the reason for the 2/3 slope was because brain volume scaled as a surface area function. Jerison’s data showed similar slopes for two groups of measurements. One group was for the “higher vertebrates” (mammals and birds), while the other group covered the “lower vertebrates” (fish, amphibians, and reptiles). The two groups were separated by a tenfold difference in absolute brain size. However, in the graph he shows, Jerison admits to fitting a line with 2/3 slope into both the “higher” and “lower” vertebrate polygons, rather than performing a regression analysis on the data points.
The metabolic model
More recent authors have said that the slope of the brain/body line is 3/4 and that it reflects metabolism instead. According to this model, the reason for the tenfold difference in the absolute brain size for “higher” and “lower” vertebrates is their difference in metabolism, warm-blooded vs. cold-blooded. Resting metabolic rates for mammals are usually reported as four to ten times those of amphibians and reptiles of the same body size.
Dinosaur brain/body size
There is good data on brain size for many types of dinosaurs. However, there is not enough data over a wide enough range of body sizes to determine whether the regression line has a slope of 2/3 or 3/4. In view of the possibility that metabolism may have varied considerably among different dinosaur groups, it is perhaps unwise to attempt to construct a model to cover dinosaurs as a whole, anyway.
Our information about the sizes of dinosaur brains comes from natural and artificial casts of the brain cavities of fossils. Natural sedimentary endocasts are occasionally preserved. More commonly, fossil skulls are sectioned and cast with latex in the lab. The volume of the braincase is then determined by graphic double integration or by water immersion.
For mammals, the volume of the endocast closely reflects the volume of the brain in life. This is also taken to be the case for birds, pterosaurs, and sauropod dinosaurs, because of well-preserved surface features on the endocast. This is not the case for living reptiles. Their brains usually only fill about half the volume of the endocranial cavity. Data on theropod and ornithischian dinosaurs used in this analysis has been adjusted for this.
It is commonly said that dinosaurs had unusually small brains and must have been very stupid. Ostrom and Jerison say dinosaur brains were appropriate for reptiles once the negative allometric (scaling) relationship is taken into account. Data from dinosaurs added to the brain size/body size graph only extend the “lower vertebrate” polygon into larger body sizes without distorting its shape.
What can be gleaned from fitting a regression line to the data is a measure called the “encephalization quotient” or EQ. This is the ratio of a given dinosaur’s brain size to the value predicted by the regression line. Living crocodilians have an EQ of 1.
If, indeed, brain/body size reflects metabolism, we shoud expect those archosaurs who have EQs greater than 1 to have been the ones with the highest metabolisms. Perhaps not coincidentally, the dinosaurs with the highest EQs also had the highest speed and agility as inferred from morphological characters. The ones with EQs less than 1 might have had the lowest metabolisms.
INFERRED METABOLIC RATES OF ARCHOSAURS
Now I will try to pull all the evidence together and draw some conclusions about warm- or cold-bloodedness for various types of extinct archosaurs.
Large sauropods
This group contains the familiar Apatosaurus (brontosaurus) type forms like what is seen on the Sinclair Oil signs. They were huge dinosaurs with big bodies, long necks, and small heads. They also have the lowest EQs of any of the dinosaurs studied (.25 or less). Some say they couldn’t have eaten enough with their peglike teeth to fuel an endothermic metabolism. Others say their teeth functioned only for cropping vegetation, not chewing it up. Many dinosaurs used gizzard stones to grind up their food, like birds do. Such gastroliths are sometimes found inside the rib cage of fossil skeletons. Therefore, I do not believe the weak teeth argument holds much water. However, I do think the evidence for warm-bloodedness is unconvincing for the large sauropods. My guess is that they were inertial homeotherms.
Large ornithischians
This group contains the ankylosaurs, stegosaurs, ceratopsians, and ornithopods. Based on low EQs and locomotor information, I consider the stegosaurs and ankylosaurs (EQs between .5 and 1.0) to have been good candidates for inertial homeothermy as well.
Ceratopsians and ornithopods have EQs from 0.6 to 1.5. These dinosaurs had highly developed shearing and grinding teeth, like might be useful for dinosaurs who needed to fuel a higher metabolism. This group includes the Arctic hadrosaurs. Although the evidence is somewhat equivocal, my guess is that some of this group of dinosaurs were at least rudimentary endotherms. Horner’s suggestion of a high metabolism for hadrosaurs when young, decreasing to inertial homeothermy as ontogeny progressed, is appealing.
Pterosaurs
Since pterosaurs were furry and capable of sustained flight, there is no question in my mind but what they were warm-blooded. Their EQs plot right around 1, indicating that this is probably the lower limit for a warm-blooded animal.
Theropods
This group includes the large carnosaurs and the smaller coelurosaurs. They all have EQs greater than 1, and in some cases (Archaeopteryx), above those enclosed by the reptilian polygon. Oviraptor and Stenonychosaurus have EQs well within the bird range. They were the most agile and active dinosaurs. Theirs is the lineage which gave rise to birds. I consider it unequivocal that advanced coelurosaurs were endotherms. Possibly all theropods were.
Birds as archosaurs
Hopefully I will not ruffle any feathers by declaring birds to be warm-blooded. Their high metabolism was bequeathed to them by their dinosaurian ancestors. Endothermy and insulation may have been the decisive factors in their escaping the wave of Late Cretaceous extinctions when their cousins could not. Dingus and Rowe, in their book The Mistaken Extinction, state that birds are “card-carrying, avialian, maniraptoran, coelurosaurian, tetanurine, theropod, saurischian dinosaurs, and don’t you forget it!” They would include birds with other dinosaurs in a cladistic classification which uses groups of hierarchical rank that do not conform to the traditional Linnaean system of Class, Order, etc. However you want to look at it, the conclusion is inescapable that dinosaurs didn’t really become extinct at the end of the Cretaceous; only the large ones did. That’s comforting news for those of us who have been dinophiles since our preschool years.
CONCLUSIONS
Indisputable evidence does not exist for warm-bloodedness in all types of dinosaurs. However, it is highly unlikely that all dinosaurs were cold-blooded, either. Morphological and postural evidence, bone histology, ecological information, and brain/body size relationships indicate that we cannot make sweeping generalizations about dinosaurian metabolism. Most likely it varied between groups. Dinosaurs spanned the spectrum from inertial homeotherms to active, endothermic birds.
Courtesy: Essay By Lynne M Clos in Fossilnews.com
 Posted in
Posted in  Tags:
Tags: 