

 March 11th, 2013
March 11th, 2013  Riffin
Riffin I. Various Species Concepts
A. Determining whether two plants are members of the same species has intriqued and perplexed biologists for a very long time. Indeed, some question the reality of species from a philosophical standpoint, such as nominalism and realism (see Wikipedia for discussion of this as well as definitions of species). Darwin said that species did not matter much and basically expressed the idea that they are whatever a competent taxonomist says they are. If we follow this idea, you can all go home now – and make sure you become competent when we ask about your species concept! In 1984 Warren H. Wagner Jr. (biography) came up with the following definition of species that helps to set the stage for examining what criteria have been used by scientists to define species.
B. Biological Species Concept (= genetic species concept, isolation species concept, etc.).
1. First used by the entomologist Jordan (1896) and later by Mayr (1969) – “a group of interbreeding populations which are reproductively isolated from other groups”. Mayr later (1982) redefined species as “a reproductive comunity of populations (reproductively isolated from others) that occupies a specific niche in nature.” The genetic species concept aimed to use measured genetic difference (distance) between populations. Problem with B.S.C. w/ plants:
a. Gene flow. Just because there is gene flow does not mean that the plants are not distinct lineages. Numerous examples of hybridization between species exist, but these parental species remain distinct in nature.
b. Asexual species. What about apomictic species? Should each individual be considered a species because it does not interbreed with other individuals?
2. Reproductive isolation was generally associated with allopatry. The following chart from Mayr illustrates the relationships between these concepts.
| Individuals Are | Not Reproductively Isolated |
Reproductively
Isolated |
| Identical in morphology and SYMPATRIC |
Same Population | Sibling Species |
| Identical in morphology and ALLOPATRIC |
Same Subspecies | Sibling Species |
| Different in morphology and SYMPATRIC |
Variant Individuals of the same population |
Separate Species |
| Different in morphology and ALLOPATRIC |
Separate subspecies |
Separate Species |
D. Phenetic Species Concept. The criteria used to define species involved gaps in character variation. These ideas were advanced by pheneticists suchas Sokal and Sneath in the 1960s and 70s. They recognized distinct phenetic clusters obtained following numerical analyses (UPGMA, PCA, etc.). One problem is determining at what level one should establish as being a species level difference, i.e. where is the phenon line set?
E. Evolutionary Species Concept. The criterion was to use common evolutionary fate through time (Wiley 1978). According to Simpson (1951) “a lineage evolving separately from others with its own unitary evolutionary role and tendencies”.
F. Paleontological Species Concept (= Successional, Chronospecies). This obviously applies to groups with a fossil record, which makes it untenable for many groups without such data. Also, fossil material presents its own unique set of problems – i.e. incomplete material, inability to conduct crosses, measure gene flow, etc.
G. Phylogenetic Species Concept (= Cladistic or Apomorphic Species Concept).
4. Olmstead (1995) proposed using the terms “apospecies” for those that possess a uniquely derived character and “plesiospecies” for those that lack such characters (Figure from his paper). These are terms that are applied once one has hypotheses of relationships (i.e. trees). These terms are similar to “cladospecies” and “paraspecies” of Ackery and Vane-Wright (1984).
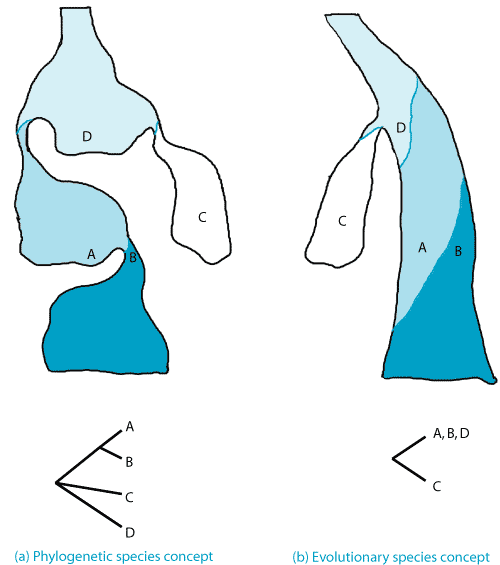
H. Diagnosabililty Species Concept. Proposed by Nixon and Wheeler (1990), it defined a phylogenetic species as “the smallest aggregation of populations (sexual) or lineages (asexual) diagnosable by a unique combination of [fixed] character states in comparable individuals.” But is a small, diagnosable change found in a series of populations (say a single base change on DNA) sufficient to recognize these populations as different species? Also, how much sampling is required to determine whether any character is truly fixed in a species?
I. Exclusivity Species Concept (= Geneological, Coalescent). Proposed by Baum and Shaw (1995), states that members of a group must be “more closely related to one another than to any organisms outside the group”. Exclusive groups that contain no less inclusive groups are Geneological Species. Exclusivity is determined by gene coalescence, i.e. if the genes of one species can be traced historically to a common ancestor and are more related to each other than to genes of another species. Sets of organisms (populations) for which monophyly cannot be demonstrated are termed “metaspecies”.
II. Plant species – some other topics
A. Are plant species really different from animals? Rieseberg et al. (2006) analyzed phenetic and/or crossing data from over 400 genera of plants and animals. Although discrete phenotypic clusters existed in > 80% of the genera, there was poor (< 60%) correspondence of taxonomic species to these clusters AND this was not different for plants or animals. 75% of phenotypic clusters in plants corresponded to reproductively isolated lineages (postmating isolation). Plant species turned out to be more likely than animal species to represent reproductively independent lineages.
B. Six paper in Systematic Botany (volume 20 from 1995) were devoted to the species problem in plants. They were by Davis, Baum and Donoghue, Doyle, Luckow, McDade and Olmstead (see citations below).
References
Ackery, R. R., and R. I. Vane-Wright. 1984. Milkweed butterflies: their cladistics and biology. Cornell University Press, Ithaca, NY.
Baum, D. A., and K. L. Shaw. 1995. Genealogical perspectives on the species problem in P. C. Hoch and A. G. Stephenson, eds. Monographs in Systematic Botany. Missouri Botanical Garden, St. Louis.
Baum, D. A., and M. J. Donoghue. 1995. Choosing among alternative “Phylogenetic” species concepts. Systematic Botany 20: 560-573. JSTOR.
Cronquist, A. 1988. The evolution and classification of flowering plants. New York Botanical Garden, Bronx, New York.
Davis, J. I. 1995. Species concepts and phylogenetic analysis – introduction. Systematic Botany 20: 555-559. JSTOR.
Donohue, M. J. 1985. A critique of the biological species concept and recommendations for a phylogenetic alternative. Bryologist 88: 172-181. JSTOR.
Doyle, J. J. 1995. The irrelevance of allele tree topologies for species delimitation, and a non-topological alternative. Systematic Botany 20: 574-588. JSTOR.
Jordan, K. 1896. On mechanical selection and other problems. Novit. Zool. 3: 426-525.
Luckow, M. 1995. Species concepts: assumptions, methods, and applications. Systematic Botany 20: 589-605. JSTOR.
Mayr, E. 1969. Principles of systematic zoology. McGraw-Hill, New York.
Mayr, E. 1982. The Growth of Biological Thought: Diversity, Evolution and Inheritance. Harvard University Press, Cambridge.
McDade, L. 1995. Species concepts and problems in practice: insight from botanical monographs. Systematic Botany 20: 606-622. JSTOR.
Mishler, B. D. 1985. The morphological, developmental, and phylogenetic basis of species concepts in bryophytes. Bryologist 88: 207-214. JSTOR.
Nixon, K. C., and Q. D. Wheeler. 1990. An amplification of the phylogenetic species concept. Cladistics 6: 211-223.
Olmstead, R. 1995. Species concepts and plesiomorphic species. Systematic Botany 20: 623-630. JSTOR.
Rieseberg, L. H. 1992. The genetic basis of morphological differences between plant species. International Journal of Plant Sciences, 153:5-6. JSTOR.
Rieseberg, L. H., and L. Brouillet. 1994. Are many plant species paraphyletic? Taxon 43: 21-32. JSTOR.
Rieseberg, L. H., T. E. Wood, and E. J. Baack. 2006. The nature of plant species. Nature 440: 524-527. HERE.
Simpson, G. G. 1951. The species concept. Evolution 5: 285-298. JSTOR.
Sluys, R. 1991. Species concepts, process analysis, and the hierarchy of nature. Experientia (Basel) 47: 1162-1170.
Wagner, W. H. 1984. A comparison of taxonomic methods in biosystematics. Pages 643-654 in W. F. Grant, ed. Plant Biosystematics. Academic Press, Toronto.
Wiley, E. O. 1978. The evolutionary species concept reconsidered. Systematic Zoology 24: 17-26. JSTOR.
Source: SIUC / College of Science / Plant Biology / PLB 479 / Lecture PLB479/ Species Concepts
 Posted in
Posted in  Tags:
Tags: 
{kind=link}