

 August 23rd, 2016
August 23rd, 2016  Riffin
Riffin @WFS News,WFS,World Fossil Society,Riffin T Sajeev,Russel T Sajeev
Beetles with Orchid Pollinaria in Dominican and Mexican Amber
Orchids are extraordinary plants that have evolved the strategy of dispersing their pollen in little sacs called pollinia. Pollinia are normally attached by supports (caudicles) to adhesive pads (viscidia) that stick to various body parts of the pollinator. The entire pollination unit is called a pollinarium. Since pollen dispensed in pollinia is not available as food to most insects (van der Cingel 1995), orchids supply nectar or have evolved ingenious patterns of mimicry and deceit to attract potential pollinators (Endress 1994).

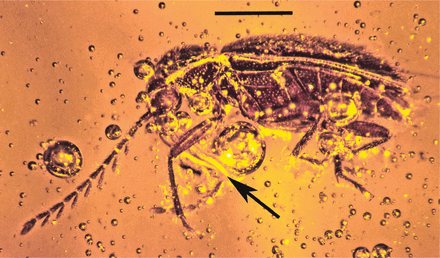
Dominican amber cryptorhynchid weevil with attached pollinarium (arrow) of Cylindrocites browni gen. n., sp. n. Floral remains in background. Scale = 3.0 mm. @WFS News
Beetle associations with orchids are diverse. Beetles may visit orchid flowers simply for nectar, or at the same time acquire pollinaria and serve as pollinators. Beetles can also feed and raise their brood on orchid floral and vegetative parts. Some orchids depend on various Coleoptera for pollination and have produced “cantharophilous” flowers especially attractive to beetles (Endress 1994, Kalshoven and van der Laan 1981).

Lateral view of weevil with attached pollinarium of Cylindrocites browni gen. n., sp. n. Arrow shows viscidium. Scale = 0.8 mm. @WFS News
The present study provides the first fossil evidence of beetles serving as pollinators of orchids. The two examples consist of a cryptorhynchine weevil (Coleoptera: Curculionidae) in Dominican amber with an orchid pollinaria attached to its thorax and a ptilodactyline beetle (Coleoptera: Ptilodactylidae) in Mexican amber with a pollinarium adhering to its mouthparts. These pollinaria are described below and associations between beetles and orchids are reviewed.

Lateral view of pollinarium of Cylindrocites browni gen. n., sp. n. attached to weevil. V = viscidium. Top arrow shows mucoid connection between viscidium and weevil. Bar = 0.4 mm. Insert shows pollen tetrads (arrow) inside pollinium. Scale = 60 μm.@WFS News
Materials and methods
The Dominican amber specimen was obtained from mines located in the Cordillera Septentrional of the Dominican Republic. These mines are in the Altimira facies of the El Mamey formation, which is shale-sandstone interspersed with a conglomerate of well-rounded pebbles (Eberle et al. 1980). Ages range from 20–15 Ma (Iturralde-Vinent and MacPhee 1996) and 20–23 million years (Baroni-Urbani and Saunders 1980) based on foraminifera counts and 45–30 Ma based on coccoliths (Cépek in Schlee 1990). These are considered minimum dates because they are based on microfossils in the strata containing the amber. Most of the amber is secondarily deposited in turbiditic sandstones of the Upper Eocene to Lower Miocene Mamey Group (Draper et al. 1994).
Mexican amber ptilodactyline beetle with attached pollinium of Annulites mexicana gen. n., sp. n. (arrow). Scale = 0.4 mm.@WFS News
The Mexican amber specimen originated from the Simojovel area of Chiapas, Mexico. Amber in this region occurs in lignitic beds among sequences of primarily marine calcareous sandstones and silt. The amber occurs in association with the Balumtun Sandstone formation of the early Miocene and the La Quinta formation of the Late Oligocene (Poinar 1992). These formations have been assigned radiometric ages from 22.5 to 26 million years (Berggren and Van Couvering 1974).

Pollinium (arrow) of Annulites mexicana gen. n., sp. n. attached to the mouthparts of a ptilodactyline beetle in Mexican amber. Scale = 0.3 mm.
Observations and photographs were made with a Nikon SMZ-10 R stereoscopic microscope and Nikon Optiphot compound microscope with magnification up to 800×. Helicon Focus Pro X64 was used to stack photos for better clarity and depth of field.
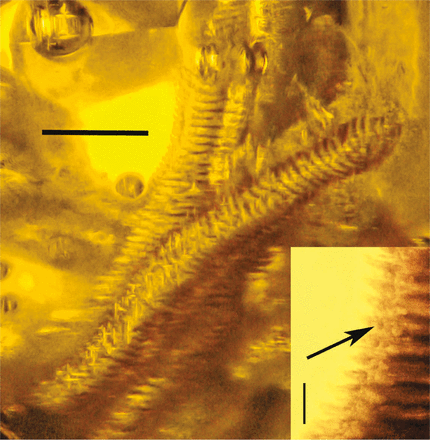
Detail of annulated pollinia of Annulites mexicana gen. n., sp. n. Scale = 0.9 mm. Insert shows pollen tetrads (arrow) between segments. Scale = 80 μm.@WFS News
Citation: DOI: http://dx.doi.org/10.1093/ae/tmw055 172-177 First published online: 22 August 2016
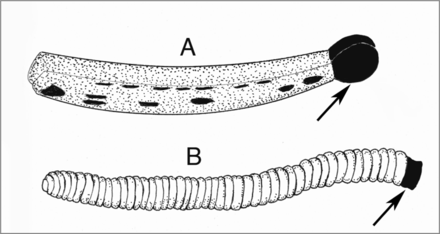
Outlines of pollinaria of Cylindrocites browni gen. n., sp. n. (A) and Annulites mexicana gen. n., sp. n. (B). Arrows show viscidia.@WFS News
Key: @WFS News,WFS,World Fossil Society,Riffin T Sajeev,Russel T Sajeev
 Posted in
Posted in  Tags:
Tags: 
This is the perfect site for everyone who really wants to understand this topic. You understand a whole lot its almost hard to argue with you (not that I personally will need toÖHaHa). You certainly put a new spin on a subject which has been discussed for many years. Excellent stuff, just great!