May 19th, 2013
May 19th, 2013  Riffin
Riffin A scientist has discovered an ancient extinct creature with ‘scissor hand-like’ claws in fossil records and has named it in honour of his favourite movie star.
The 505-million-year-old fossil called Kooteninchela deppi (pronounced Koo-ten-ee-che-la depp-eye), which is a distant ancestor of lobsters and scorpions, was named after the actor Johnny Depp for his starring role as Edward Scissorhands — a movie about an artificial man named Edward, an unfinished creation, who has scissors for hands.
Kooteninchela deppi is helping researchers to piece together more information about life on Earth during the Cambrian period when nearly all modern animal types emerged.
David Legg, who carried out the research as part of his PhD in the Department of Earth Science and Engineering at Imperial College London, says:
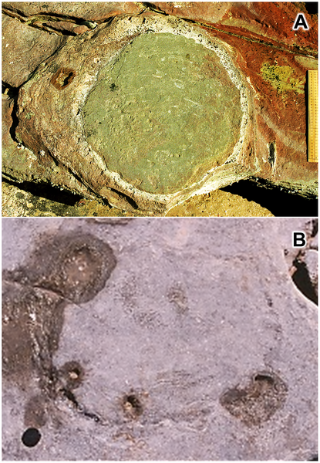
“When I first saw the pair of isolated claws in the fossil records of this species I could not help but think of Edward Scissorhands. Even the genus name, Kootenichela, includes the reference to this film as ‘chela’ is Latin for claws or scissors. In truth, I am also a bit of a Depp fan and so what better way to honour the man than to immortalise him as an ancient creature that once roamed the sea?”
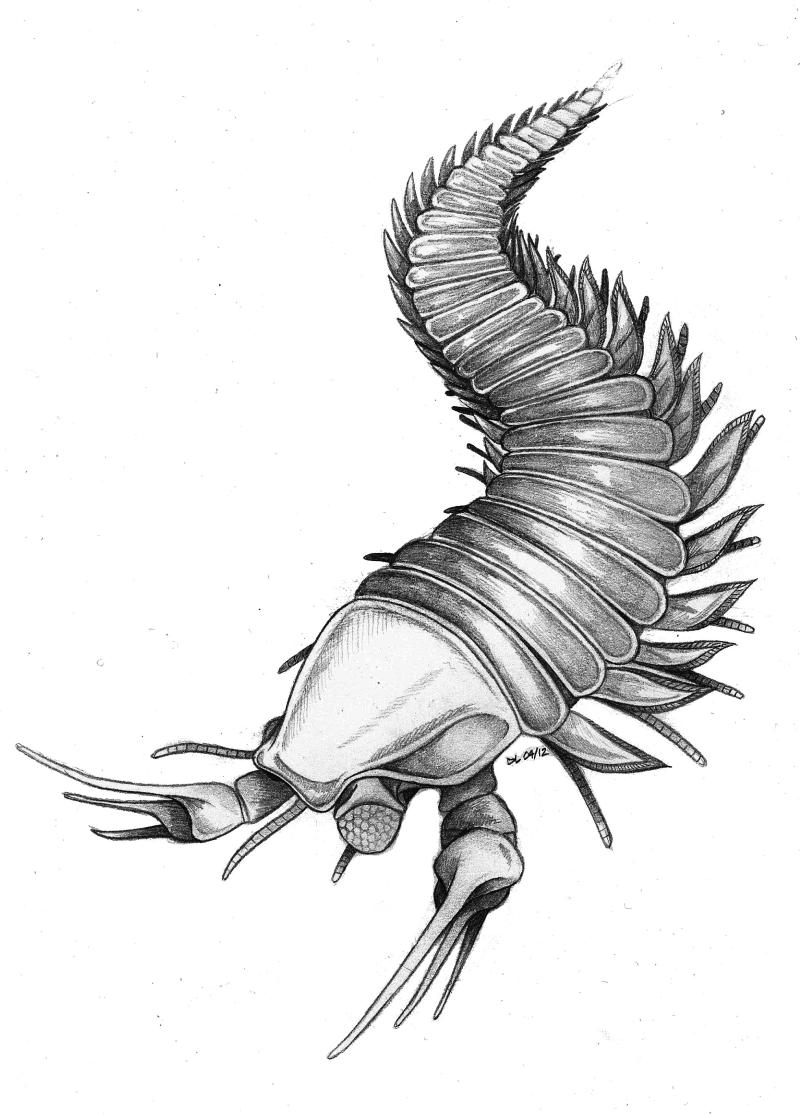
Kootenichela reconstruction. (Credit: Image courtesy of Imperial College London)
Kooteninchela deppi lived in very shallow seas, similar to modern coastal environments, off the cost of British Columbia in Canada, which was situated much closer to the equator 500 million years ago. The sea temperature would have been much hotter than it is today and although coral reefs had not yet been established, Kooteninchela deppi would have lived in a similar environment consisting of sponges.
The researcher believes that Kooteninchela deppi would have been a hunter or scavenger. Its large Edward Scissorhands-like claws with their elongated spines may have been used to capture prey, or they could have helped it to probe the sea floor looking for sea creatures hiding in sediment.
Kooteninchela deppi was approximately four centimetres long with an elongated trunk for a body and millipede-like legs, which it used to scuttle along the sea floor with the occasional short swim.
It also had large eyes composed of many lenses like the compound eyes of a fly. They were positioned on top of movable stalks called peduncles to help it more easily search for food and look out for predators.
The researcher discovered that Kooteninchela deppi belongs to a group known as the ‘great-appendage’ arthropods, or megacheirans, which refers to the enlarged pincer-like frontal claws that they share. The ‘great-appendage’ arthropods are an early relation of arthropods, which includes spiders, scorpions, centipedes, millipedes, insects and crabs.
David Legg adds: “Just imagine it: the prawns covered in mayonnaise in your sandwich, the spider climbing up your wall and even the fly that has been banging into your window and annoyingly flying into your face are all descendants of Kooteninchela deppi. Current estimates indicate that there are more than one million known insects and potentially 10 million more yet to be categorised, which potentially means that Kooteninchela Deppi has a huge family tree.”
In the future, David Legg intends to further his research and study fossilised creatures from the Ordovician, the geological period that saw the largest increase in diversity of species on the planet. He hopes to understand why this happened in order to learn more about the current diversity of species on Earth.
The research was published in the Journal of Palaeontology 2 May 2013.


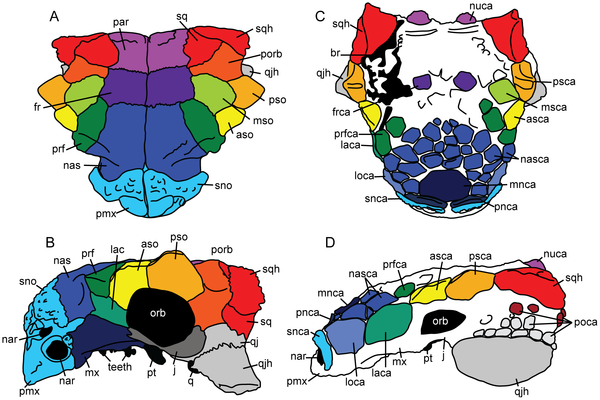
![Cretaceous sauropod tracks and their potential rack-makers. A, silhouette of Diamantinasaurus, a titanosaur or related sauropod from the Winton Formation (Albian-Cenomanian) of Queensland (after Hocknull et al. [40]); scale bar indicates 1 metre. B, silhouette of Brachiosaurus (after Farlow [19]); undescribed skeletal fragments of a similar sauropod are also known to occur in the Rolling Downs Group of Queensland; scale bar indicates 1 metre. C, right manus-pes couple (at right) and D, part of a trackway (at left), of Brontopodus birdi, a distinctive form of sauropod track from the Trinity Group (Early Cretaceous, Comanchean) of Texas and Arkansas; after Farlow et al. [38]); long suspected to be the track of the contemporary brachiosaur Pleurocoelus, but more recently attributed [58] to Paluxysaurus, a relative of Brachiosaurus; length of the pes print ranges from 40–50 cm to more than 100 cm. E, a sample of sauropod tracks from the Broome Sandstone, Western Australia, to illustrate their diversity in size and shape; three isolated pes prints (at left) and three manus-pes couple (at right) are shown at uniform scale; scale bar (extreme left) is 1 metre. doi:10.1371/journal.pone.0036208.g002](http://worldfossilsociety.org/wp-content/uploads/2013/05/121219174150-large2.png)



![Temporal and geographical sampling, and terminology on conodont elements. (A) Timescale, composite section along the end Frasnian and the Famennian and stratigraphic log of the studied sections. Absolute ages after [24] and conodont zones after [77]. Note that the postera and expansa zones are not sampled. Along the stratigraphic log of each section, dots represent the sampled levels. Abbreviations: E = Early, M = Middle, L = Late. (B) Paleogeographic map [19] of the Famennian. Circles = location of the French (blue circle) and German (red circle) sections contributing to the composite section. Black dots = location of the sections that delivered additional sampling of Palmatolepis (Palmatolepis) linguiformis. (C) Illustration of Palmatolepis platform elements, with the terminology of the morphological features used in taxonomy. To the left a specimen of Pa. (Palmatolepis) rugosa (trachytera zone). To the right a specimen of Pa. (Manticolepis) rotunda (rhenana zone). doi:10.1371/journal.pone.0036230.g001](http://worldfossilsociety.org/wp-content/uploads/2013/05/121219174150-large1.png)