











 September 10th, 2019
September 10th, 2019  Riffin
Riffin @WFS,World Fossil Society,Riffin T Sajeev,Russel T Sajeev
Research published in Scientific Reports describes Clevosaurus hadroprodon, a new reptile species from Rio Grande do Sul state in southern Brazil. Its fossils remains — jaws and associated skull bones — were collected from Triassic rocks (c. 237-228 million-years old) making it the oldest known fossil of its kind in Gondwana, the southern supercontinent that would eventually become Africa, Antarctica, Australia, India, and South America.

Clevosaurus hadroprodon was a small animal, similar in size with common house geckos. It belongs to the Sphenodontia, a group of lepidosaurs (which also includes snakes, lizards and amphisbaenians), that was very diverse and widespread during the Mesozoic era (the “Age of Dinosaurs”), but today has only one remaining living species in New Zealand. Clevosaurus hadroprodon is the oldest member of the Clevosauridae, a group of small sphenodonts that were the first globally distributed lepidosaurs with fossils from the Late Triassic and Early Jurassic of North America, Europe, Asia, Africa and South America.
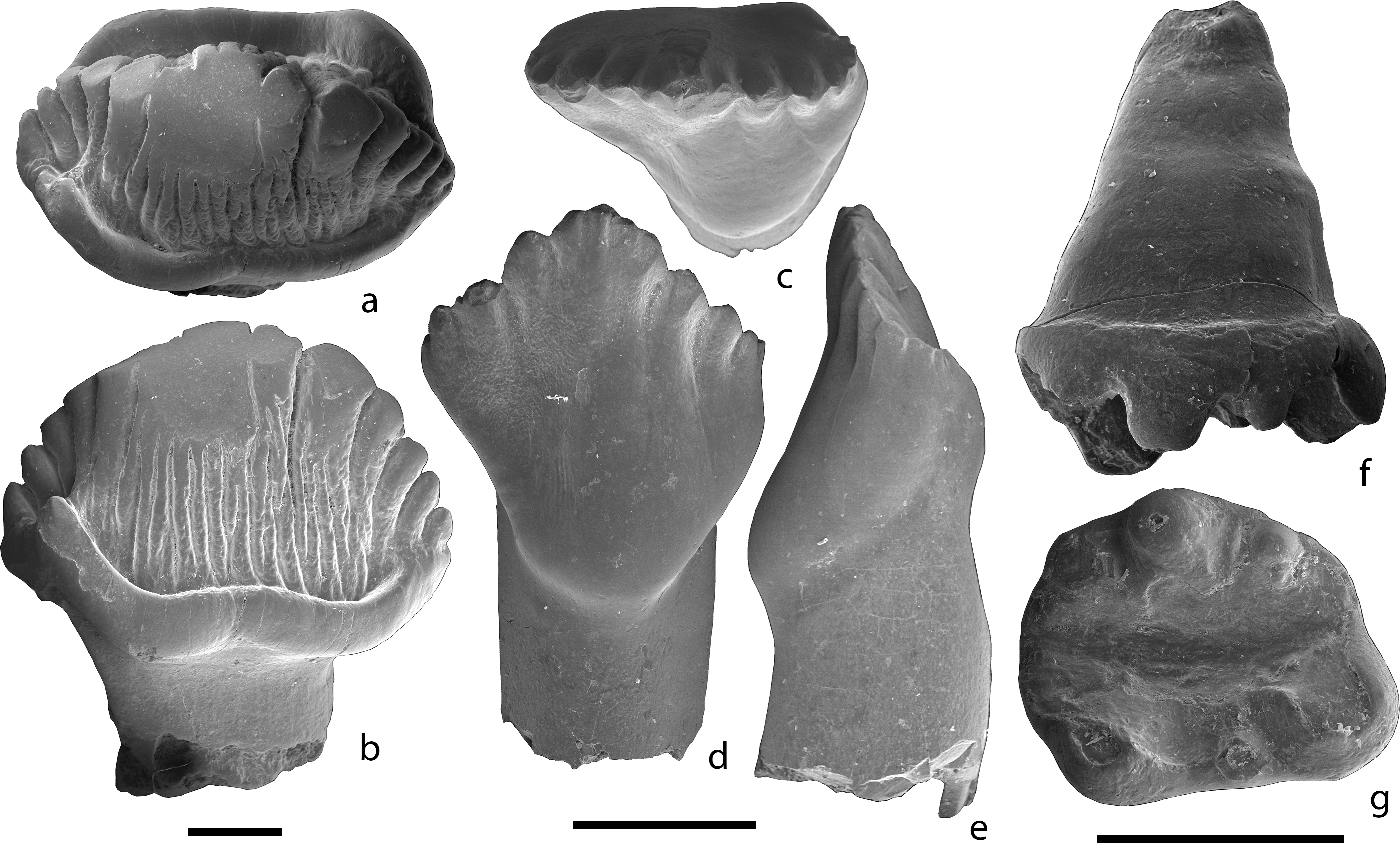
The dentition of Clevosaurus hadroprodon is an unexpected mix of primitive and derived teeth. It is the oldest occurrence of the typical fully acrodont dentition (teeth fused to the top of the jaw bones) of sphenodontians, but most of its teeth are relatively simple and blade-like, which differs from other, only slightly younger Clevosaurus species that possess well-developed medial-posteromedial (side-to-side) expansions of the teeth for complex grinding. “However, Clevosaurus hadroprodon also possess a large, blunt, tusk-like tooth in the first tooth position of the both premaxilla (upper jaw) and of dentary (lower jaw). This feature is typically observed only in later sphenodontian lineages” says Annie Schmaltz Hsiou, Associate Professor at the University of São Paulo and head of the study. The name “hadroprodon” is Greek for “larger first tooth” in reference to these tusk-like teeth.
“Clevosaurus hadroprodon is an important discovery because it combines a relatively primitive sphenodontian-type tooth row with the presence of massive tusk-like teeth that were possibly not for feeding, but rather used for mate competition or defense. If correct, this means that non-feeding dental specializations predated changes in the sphenodontian dentition related to feeding strategies. This is a very exciting discovery.” says co-author Randall Nydam, Professor at Midwestern University (US).
In addition to its unique dentition, the authors stress that Clevosaurus hadroprodon also adds to the growing evidence that the early diversification of sphenodontians occurred in the widely separated regions of Gondwana destined to become South American and India. This illustrates the importance of the role of the Gondwanan lepidosaur fauna in our growing understanding of the earliest stages of sphenodontian evolution and the global biogeographic distribution of lepidosaurs.
- Annie S. Hsiou, Randall L. Nydam, Tiago R. Simões, Flávio A. Pretto, Silvio Onary, Agustín G. Martinelli, Alexandre Liparini, Paulo R. Romo de Vivar Martínez, Marina B. Soares, Cesar L. Schultz, Michael W. Caldwell. A New Clevosaurid from the Triassic (Carnian) of Brazil and the Rise of Sphenodontians in Gondwana. Scientific Reports, 2019; 9 (1) DOI: 10.1038/s41598-019-48297-9





![Fig. 1 ?Crossvallia waiparensis, sp. nov. from the Waipara Greensand in New Zealand. A‒L, overview of the leg bones preserved in the holotype (CM 2018.23.9) and M‒O, tentatively referred proximal end of a left humerus (CM 2016.158.3). A‒C, Left tibiotarsus in cranial (A), caudal (B) and lateral (C) view. D‒F, Right tibiotarsus in cranial (D), caudal (E) and lateral (F) view. G, H, Distal end of left femur in cranial (G) and caudal (H) view. I‒K, Right tarsometatarsus in cranial (I), plantar (J) and distal (K) view. L, Pedal phalanx. M‒O, Tentatively referred proximal end of left humerus in caudal (M), ventral (N) and cranial (O) view. Abbreviations: cms, crista musculi supracoracoidei; fvd, foramen vasculare distale; fvp, foramen vasculare proximale; pst, pons supratendineus; stv, sulcus transversus. Scale bar = 50 mm. [Colour online].](http://www.worldfossilsociety.org/wp-content/uploads/2019/08/190220161034-ancient-finds-exlarge-169-2.jpg)
![Fig. 2 ?Crossvallia waiparensis, sp. nov. from the Waipara Greensand, bones of the holotype (CM 2018.23.9) in comparison with the extant Emperor penguin (Aptenodytes forsteri) and other penguins from the Paleocene of New Zealand. A‒C, ?C. waiparensis (holotype), distal end of left femur in cranial (A), caudal (B), and distal (C) view. D, Kumimanu biceae (holotype, NMNZ S.45877), distal end of right femur in distal view (some surrounding matrix digitally removed). E‒J, Sequiwaimanu rosieae (holotype, CM 2016.6.1), distal end of right femur in cranial (E, H), caudal (F, I) and distal (G, J) view; in H‒J, the bones are mirrored and brought to the same size as the ?C. waiparensis femur. K, Tibiotarsus of Aptenodytes forsteri in cranial view (left side, mirrored). L, ?C. waiparensis (holotype), right tibiotarsus in cranial view. M, Distal end of right tibiotarsus of Waimanu manneringi (holotype, CM zfa 35) in cranial view. N, O, Distal end of left tibiotarsus of S. rosieae (holotype, CM 2016.6.1) in cranial view; in O, the bone is mirrored and brought to the same size as the ?C. waiparensis tibiotarsus. P, Q, Partial right tibiotarsus of K. biceae (holotype, NMNZ S.45877) in cranial view; in P, the condylus medialis was digitally brought in its presumed original position and adhering bone fragments and matrix were digitally removed. R, S, ?C. waiparensis (holotype), right tarsometatarsus in plantar (R) and dorsal (S) view. T, U, Partial left tarsometatarsus of an unnamed very large penguin from the Waipara Greensand (CM 2016.158.1) in plantar (T) and dorsal (U) view. V, W, Right tarsometatarsus of W. manneringi (holotype, CM zfa 35) in plantar (V) and dorsal (W) view. X, Y, Right tarsometatarsus of Muriwaimanu tuatahi (CM zfa 34) in plantar (X) and dorsal (Y) view. Abbreviations: cdl, condylus lateralis; cdm, condylus medialis; mpr, medial projection of proximal tarsometatarsus; prj, proximal projection of crista patellaris; sfb, semicondylus fibularis; spt, sulcus patellaris; stf, semicondylus tibiofibularis. Scale bars = 50 mm; same scale for all figure panels except H‒J and O. [Colour online].](http://www.worldfossilsociety.org/wp-content/uploads/2019/08/190220161034-ancient-finds-exlarge-169-3.jpg)
![Fig. 3 A‒C, Holotype (CM 2018.23.9) and D‒F, tentatively referred proximal humerus (CM 2016.158.3) of ?Crossvallia waiparensis, sp. nov. from the Waipara Greensand in New Zealand, in comparison with G‒L, the holotype of Crossvallia unienwillia (MLP 00-I-10-1) from the Thanetian of Antarctica. A, B, Distal end of left femur in cranial (A) and caudal (B) view. C, Distal end of right tibiotarsus in cranial view; the dotted line indicates the sulcus extensorius. D‒F, Proximal end of tentatively referred left humerus in caudal (D), ventral (E) and cranial (F) view. G, H, Distal end of right femur in cranial (G) and caudal (H) view. I, Distal end of right tibiotarsus in cranial view; the dotted line indicates the sulcus extensorius. J‒L, Proximal end of left humerus in caudal (J), ventral (K) and cranial (L) view; the dotted lines indicate the broken portion of the bone. Scale bar = 50 mm. [Colour online].](http://www.worldfossilsociety.org/wp-content/uploads/2019/08/190220161034-ancient-finds-exlarge-169-4.jpg)
