May 25th, 2017
May 25th, 2017  Riffin
Riffin @WFS,World Fossil Society,Riffin T Sajeev,Russel T Sajeev
Citation: Djokic, T. et al. Earliest signs of life on land preserved in ca. 3.5 Ga hot spring deposits. Nat. Commun. 8, 15263 doi: 10.1038/ncomms15263 (2017).
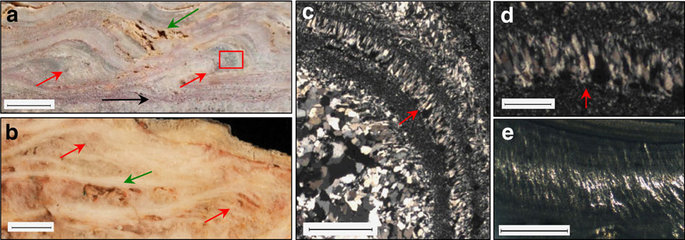
Scale bar measurements indicated. (a) Dresser terracettes (red arrows) with preserved primary porosity (green arrow) and a horizon containing Dresser stratiform geyserite (black arrow). Scale bar, 1 cm. Inset box of c. displays palisade fabric. (b) >1,800-year-old sinter terracettes (red arrows) with preserved primary porosity (green arrow) from a sinter buttress at Te Kopia, New Zealand. Scale bar, 1 cm. Micrographs in XPL of (c) Dresser palisade fabric oriented vertical to bedding (scale bar, 1 mm) and (d) close-up (scale bar, 250 μm). (e) Sinter with preserved palisade fabric, Te Kopia, New Zealand. Scale bar, 1 mm.
@WFS,World Fossil Society,Riffin T Sajeev,Russel T Sajeev
The ca. 3.48 Ga Dresser Formation, Pilbara Craton, Western Australia, is well known for hosting some of Earth’s earliest convincing evidence of life (stromatolites, fractionated sulfur/carbon isotopes, microfossils) within a dynamic, low-eruptive volcanic caldera affected by voluminous hydrothermal fluid circulation. However, missing from the caldera model were surface manifestations of the volcanic-hydrothermal system (hot springs, geysers) and their unequivocal link with life. Here we present new discoveries of hot spring deposits including geyserite, sinter terracettes and mineralized remnants of hot spring pools/vents, all of which preserve a suite of microbial biosignatures indicative of the earliest life on land. These include stromatolites, newly observed microbial palisade fabric and gas bubbles preserved in inferred mineralized, exopolymeric substance. These findings extend the known geological record of inhabited terrestrial hot springs on Earth by ∼3 billion years and offer an analogue in the search for potential fossil life in ancient Martian hot springs.
@WFS,World Fossil Society,Riffin T Sajeev,Russel T Sajeev

(a) Proximal to distal hot spring facies, with spring vent fed by subsurface hydrothermal fluids. (b) Preserved sequence of hot spring facies deposits, geographically patchy in nature, with spring vent infilled by late-stage crystallization of barite.
@WFS,World Fossil Society,Riffin T Sajeev,Russel T Sajeev

Scale bar measurements indicated. (a) High resolution gigapan image of Dresser geyserite. Inset boxes are figure parts (b,c,d). Laminae overgrowth stages; s1 and s2 represented by white dashed lines. Ferruginous material (red arrows) contains inferred gas bubbles; see Fig. 5. Scale bar, 2 mm. Micrographs in PPL (b–f). (b) Botryoidal textures display laminae onlap/offlap (red arrow), separated by siliceous equigranular troughs (white arrow) overlain by fine, planar laminae (scale bar, 1 mm). (c) Botryoidal–columnar textures overlain by planar (black dashes), slumped (red arrow) laminae. Scale bar, 1 mm. (d) Overgrowth (e1) with outward and downward facing botryoids (white arrows). Quartz (Qz) and barite (B), infill and cross-cut laminae (scale bar, 1 mm). (e) Close-up of light/dark microlaminae in Dresser geyserite. Inset box of figure part (i). Scale bar, 50 μm. (f) Modern geyserite with botryoidal microlaminae (red arrow), Geysir, Iceland. Analogous to (b). Scale bar, 1 mm. (g) Slumped laminae of<100-year-old geyserite, Geyser Valley, New Zealand. Analogous to c. Scale bar, 1 cm. (h) Pool rim overgrowth of geyserite with outward facing botryoids (arrows), Geyser Valley, New Zealand. Analogous to d. Scale bar, 2 cm. (i) SEM-EDS element maps showing light bands enriched in K–Al alternating with dark bands enriched in Ti, identified as kaolinite+illite and anatase, respectively, from Raman spectroscopy and XRD analysis; see Supplementary Figs 2–6 (scale bar, 50 μm).